脊椎动物多点起源论及两栖爬行类动物的起源
脊椎动物多点起源论及两栖爬行类动物的起源
人们公认早期脊椎动物从无脊椎动物起源的路径是单一的,最早出现的脊椎动物是鱼类中无颌类,就像现存圆口纲中的盲鳗和七鳃鳗,现生丰富多彩、千变万化的鱼类和陆生脊椎动物,都是从无颌类这根主干发展演化而来的。鱼类又分两大类,软骨鱼类与硬骨鱼类。最早的软骨鱼类出现于泥盆纪早期(距今3亿8千万年前),以后演化成了各类的鲨鱼。硬骨鱼类是最进步的鱼类,陆地脊椎动物被认为就是从硬骨鱼类演化出来的。
一、早期脊椎动物是多点起源的。
最早出现脊椎动物是鱼类中的无颌类,以后的所有脊椎动物都是从这根主干发展演化出来的这套理论,根据笔者对艾迪卡拉纪环节古虫的研究发现,是绝对错误的,事实远非如此。鱼类的起源并不是人们普遍公认的那样循单一路径起源演化,而是多点起源,并各自独立演化发展而来的,而且当时脊椎动物并不只有鱼类,还有两栖类的祖先(后面有详细论述),这时的所谓鱼类应该还不是严格意义上的鱼类。
徐淮地区的马家沟组地层上部,有对应的艾迪卡拉纪环节古虫化石证据(图1,图2,图3,),完全可以证明这一点。由于鱼类是寻各自的路径独立起源演化的,所以软骨鱼类与硬骨鱼类起源谁先谁后的争论,没有任何意义。现生的无颌类、软骨鱼类及硬骨鱼类,都是从艾迪卡拉纪不同类型的环节古虫,循各自独立路径起源演化而来的。
 图1,艾迪卡拉纪马家沟组上部向脊椎动物演化的环节古虫化石。
图1,艾迪卡拉纪马家沟组上部向脊椎动物演化的环节古虫化石。


图2,艾迪卡拉纪马家沟组上部向脊椎动物演化的环节古虫化石。


图3,艾迪卡拉纪马家沟组上部环节古虫化石,具脊椎动物的重要特征之一的咽鳃裂。
其中的软骨鱼类与硬骨鱼类都必然经历一个颌口发生的变化过程。这是生物进化历史上的一次巨大进步,捕食方式因此由被动一跃成为主动。捕食效率大大提高,生存适应性和生存机会大增,为以后脊椎动物各门类的大发展打下了坚实基础。颌口发生时间大概于艾迪卡拉纪与寒武纪之交,在华北凤山组的上部,网上见到过被咬残缺的三叶虫化石可以作为证据。
 图4,演化到寒武纪徐庄组的环节古虫,已经可以成为现在意义上的鱼类了。化石表现了由于水的干涸,鱼类集体死亡的场景。
图4,演化到寒武纪徐庄组的环节古虫,已经可以成为现在意义上的鱼类了。化石表现了由于水的干涸,鱼类集体死亡的场景。
 图5,演化到寒武纪徐庄组的环节古虫,盲鳗或七鳃鳗的祖先。
图5,演化到寒武纪徐庄组的环节古虫,盲鳗或七鳃鳗的祖先。
 图6,演化到寒武纪徐庄组,已经可以称为现在意义上的鱼类了,偶鳍还没形成。
图6,演化到寒武纪徐庄组,已经可以称为现在意义上的鱼类了,偶鳍还没形成。



笔者通过对徐淮地区广泛的早古生代地层古虫化石研究,完全可以推测出各种鱼类早期演化的轮廓(图4,图5,图6,图7)。
图4,演化到寒武纪徐庄组的环节古虫,已经可以成为现在意义上的鱼类了。化石表现了由于水的干涸,鱼类集体死亡的场景。
图5,演化到寒武纪徐庄组的环节古虫,盲鳗或七鳃鳗的祖先。
图6,演化到寒武纪徐庄组,已经可以称为现在意义上的鱼类了,偶鳍还没形成。
环节古虫向脊椎动物起源演化的开始于艾迪卡拉纪中期,相当于马家沟组地层上部,完成于馒头组上部的徐庄组。也可以说现在意义上的鱼类,是寒武纪末期诞生的。整个寒武纪期间的介于鱼类与环节古虫之间存在的早期脊椎动物,似鱼非鱼,可以称作拟鱼类,拟鱼类应包括所有由环节动物演化出来的脊椎动物(图7)。徐淮地区大量早期鱼类演化过程中的化石,可以支持并充分证明这个一点。


图7,演化到艾迪卡拉纪凤山组的环节古虫(拟鱼类),身体分节依然可辨。
从以上的这些化石证据可以看出,脊椎动物起源路径绝不是单一的,而是多点寻各自独立路径同时起源发展演化而来的。在艾迪卡拉纪马家沟组的上部,生存着诸多有着往脊椎动物方向发展潜力的环节动物,都具有脊椎动物的原始特征,咽鳃裂、背部神经管,肛後尾,……同时还有成熟的头脑,它们都是裂体腔环节动物。这与无头的文昌鱼是肠体腔动物不同,这也说明文昌鱼是不同于环节古虫的另类动物,脊椎动物不可能从无头的文昌鱼类动物起源。
二、两栖爬行类的起源。
两栖动物起源问题,始终迷雾重重,没有产生过刚性的定论,始终停留在推论假说阶段。问题出在人们的立论的根据和研究的出发点发生了错误。人们先入为主认为两栖类必须是从鱼类起源的,然后倒果为因的再去找证据,景象如雾里看花。其实,两栖类起源与不同鱼类起源一样,也是从艾迪卡拉纪不同的环节古虫独立起源进化而来的,其起源演化与各种鱼类没有任何关系,它们的起源演化是平行的(图8 )。墨西哥钝口螈就是这个进化过程中的特定阶段的孑遗生物。


图8 最新脊椎动物起源路线示意图
墨西哥钝口螈(图9),是一种比较古老的两栖类物种,原产于墨西哥,栖息在当地高山的湖泊之中。这种蝾螈最特别的地方,是它们并不会像一般两栖类一样变态进化为陆栖蝾螈,而是终其一生都著保持幼体的型态,生物学上称之为“幼体性熟”。钝口螈有着环节古虫相似的身体形态,并且有明确可分辨的与马家沟组环节古虫化石相似的身体分节,颈部两边鳃裂上各具三条羽状外鳃,这是极原始的水栖动物。
 图9 墨西哥钝口螈,来自环节古虫的残存身体分节清晰可辨(图片来源网络)
图9 墨西哥钝口螈,来自环节古虫的残存身体分节清晰可辨(图片来源网络)

从钝口螈的身体形态来看,应该是从艾迪卡拉纪环节古虫的一支(图2),独立发展演化出来的特定地质阶段的动物。称其为两栖动物,是不准确的。终生离不开水环境,是向两栖类进化的比两栖类低一个层级的水栖孑遗动物,还没进化到两栖阶段。所谓幼体性成熟(意思是曾经有过成体,莫名原因退化了)其实是误解,钝口螈就没有进化到所谓成体阶段。钝口螈是两栖类的直接祖先类型,可能是偶然因素造成其没有登上陆地,成为真正意义上的两栖类。
笔者以墨西哥钝口螈和肺鱼关键器官起源演化及身体组织的作用,来进一步说明两栖爬行类并不是从鱼类起源,而是与各种鱼类一样也是从艾迪卡拉纪环节古虫的某一支,同步独立起源演化出来的。
1、 鳃器官的产生 早期的滤食环节古虫,主要是靠身体表面呼吸的,咽鳃孔主要用来出水的。发展到了掠食阶段,演化出了颌口,由于捕食形式进步,身体活动量的增加,捕食时氧需要量骤增,外鳃的发生是必然事情。要求更快的捕食速度,于是外鳃成了累赘。因此,环节古虫摒弃了由外胚层产生外鳃,由中胚层创生出了内鳃,这部分环节古虫演化成了鱼类。对速度要求相对不高的一类环节古虫,外鳃依然保留在体外。外鳃是环节古虫发展到特定阶段的特殊身体现象,终生具外鳃的墨西哥钝口螈就是这一特定阶段的代表生物。推而广之,所有具外鳃的鱼类及两栖类,都是从环节古虫继承下来古老身体特征。从环节古虫演化而来的软骨鱼类由于一开始就性情凶猛,行动迅速,鳃孔众多,可能一开始就没有产生过外鳃。
2、
残存的体节 墨西哥钝口螈依然保存从环节古虫继承下来的身体特征和组织构造。如果不考虑其四肢,就是一条环节古虫。与环节古虫化石无论是身体分节形式,还是身体轮廓形状都高度相似。这时的环节古虫突出于体外部分翼板组织,已被身体消融吸收,残存的分节形式依然清晰可辨(图9) 。
3、 皮肤呼吸作用 是两栖类从环节古虫继承下来的固有属性,并不是人们猜测的从鱼类演化过来的次生现象。古老的生物在没有演化出来鳃之前,几乎都是体表呼吸的,现存节肢动物中这种现象依然存在。鱼类鳃非常进步,皮肤的呼吸作用完全消失了。这也是两栖类不可能从鱼类起源演化的可靠证据。强调皮肤呼吸作用,与强调皮肤保护作用是很难两全的。
4、
肢和趾的演化 往两栖类进化的环节古虫,在演化出了外鳃及上下颌之后,为了获得更大生存发展机会,又继续演化出了四肢。四肢的功用完全是为了水底的匍匐前进,隐蔽接近捕食目标,提高捕食效率。趾的发生完全是为了挂扯和抓握,使捕食活动更精确细致。两栖类在后肢演化出五趾,前肢演化出四趾后,就停止了进一步演化。至今我们看到的两栖类前肢都只具有四趾,这是进化不完全的表现。而后来的爬行类由于陆地生存需要,强调了四肢的搬运身体作用和趾的抓握作用,进而完成了前肢五趾化的演化。这样才有了中生代的爬行类动物,如统治地球长达一亿多年的身躯庞大恐龙,及现代的高级动物,如丰富多彩鸟类和哺乳类,其中的人类发展出了认识自然的高级智慧,更具划时代意义。


图10 非洲肺鱼的四肢,演化成了触须(图片来源网络)


图11 澳洲肺鱼的四肢,演化成了肉鳍(图片来源网络)
5、
鳍的产生 鳍的演化与肢的演化是循不同功用演化出来的,鳍发生目的主要是为了游泳。总鳍鱼发展出了肉鳍,肉鳍对总鳍鱼强调的是身体姿态调整能力,可以快速调整身体姿态,或许为了捕食,也或许是为了逃避天敌。肺鱼也有四只肉肢或鳍,和两栖爬行类相似,澳洲肺鱼有和总鳍鱼一样的肉鳍。笔者认为肺鱼是独立从环节古虫演化而来的,也或是与两栖类同源。肺鱼与总鳍鱼的肉鳍的进化顺序不同,肺鱼早期出现颌口的阶段为了在水中匍匐前进,演化出了肉肢,这阶段与钝口螈的演化过程一样。由于环境骤变,澳洲肺鱼又重新强调起了游泳,于是在先发生的肉肢上演化出了游泳用的鳍。非洲肺鱼的肉肢,由于环境原因,最终演化成了现在的用来探测环境用的触须,肢的功用几乎完全丧失。非洲肺鱼幼体还存在外鳃,而澳洲肺鱼幼体不存在外鳃。从以上分析可以看出,总鳍鱼是不可能登上陆地的,更不可能演化成两栖类。
6、 鳞甲的产生 鳞甲的发生也与环节古虫类动物演化出了颌口后的捕食方式的进步有关,掠食式捕食方式必须要求动物的速度和身体灵活性,于是往鱼类演化的环节古虫出了内鳃,提高了氧的摄取能力,同时抛弃了皮肤的呼吸作用,为鳞甲的发生奠定了基础。强调速度和身体灵活性必然要使裸露的皮肤冒极大受伤害的风险。因此皮肤的保护成了首要问题,鱼类的鳞甲不失时机的被大自然创造出来了。爬行类经历两栖阶段成功登陆,肺器官的演化完成,皮肤的呼吸作用完全没有必要,裸露的皮肤成了危险所在。保护皮肤成了爬行动物的进化方向,鳞甲于是应时诞生了。而两栖类中的一部分完全适应了生存环境,从其祖先继承下来的皮肤呼吸作用成了两栖类的生存法宝,没有必要放弃皮肤的呼吸作用产生鳞甲,这部分两栖类现在依然生机勃勃。
综上所述,可以看出两栖类并不是从鱼类起源的,而是从艾迪卡拉纪环节古虫中的一种,独立起源演化发展而来的。墨西哥钝口螈就是两栖类登陆前的阶段性生物,是两栖类动物从环节古虫而不是从鱼类起源的证据。两栖类的皮肤呼吸功用及羽状外鳃,是对环节古虫的原始属性的继承。四肢和不完全五趾的产生,具有巨大进步意义,前肢的四趾现象是进化不完全的表现。肺鱼有可能是两栖爬行类这条进化主干上分化比较早的一类动物,也可能是从艾迪卡拉纪环节古虫中另外一种类独立演化发展出来的。
艾迪卡拉纪环节古虫在演化成真正意义上鱼类和两栖爬行类之前,这个阶段的早期脊椎动物还不能称其为鱼类,体节正在消失的过程之中,鱼鳞还没产生,颌口刚刚演化出来,存在一个似鱼非鱼的拟鱼类阶段。
经过这个似鱼非鱼的拟鱼类阶段的早期脊椎动物,进一步演化成后来的鱼类和爬行类,必然经历过保护皮肤作用的鳞甲产生的过程。这个过程对爬行动物来讲,必然发生在两栖类阶段之后。如果鱼类的鳞甲的产生与爬行类同时,那么两栖类从鱼类起源的说法站不住脚。如果鱼类的鳞甲是更早时期产生的,那么两栖类从鱼类起源必然经历一个退鳞甲的过程,而且还需要唤醒由于鳞甲的产生丧失的皮肤呼吸作用。即使有环境的巨变所造成的严苛生存压迫,这也是不可能的事情。所以说滑皮两栖类的滑皮不是后成的,那个阶段两栖类根本就没有演化出鳞甲。滑皮两栖类可能是为了适应两栖爬行类起源于鱼类的假说,人为制造出来的概念。
参考:
http://blog.163.com/xyjq_007688/blog/static/315484272011613111439950/馒头组不是寒武纪地层,是奥陶纪地层
http://blog.163.com/xyjq_007688/blog/static/3154842720109260321582/马家沟组是一套完整的艾迪卡拉纪地层
http://blog.163.com/xyjq_007688/blog/static/31548427201071421033856/脊椎动物起源与原索动物无关——从文昌鱼谈脊椎动物起源可以休也。
http://blog.163.com/xyjq_007688/blog/static/3154842720109260321582/马家沟组是一套完整的艾迪卡拉纪地层
http://blog.163.com/xyjq_007688/blog/static/31548427201071421033856/脊椎动物起源与原索动物无关——从文昌鱼谈脊椎动物起源可以休也。
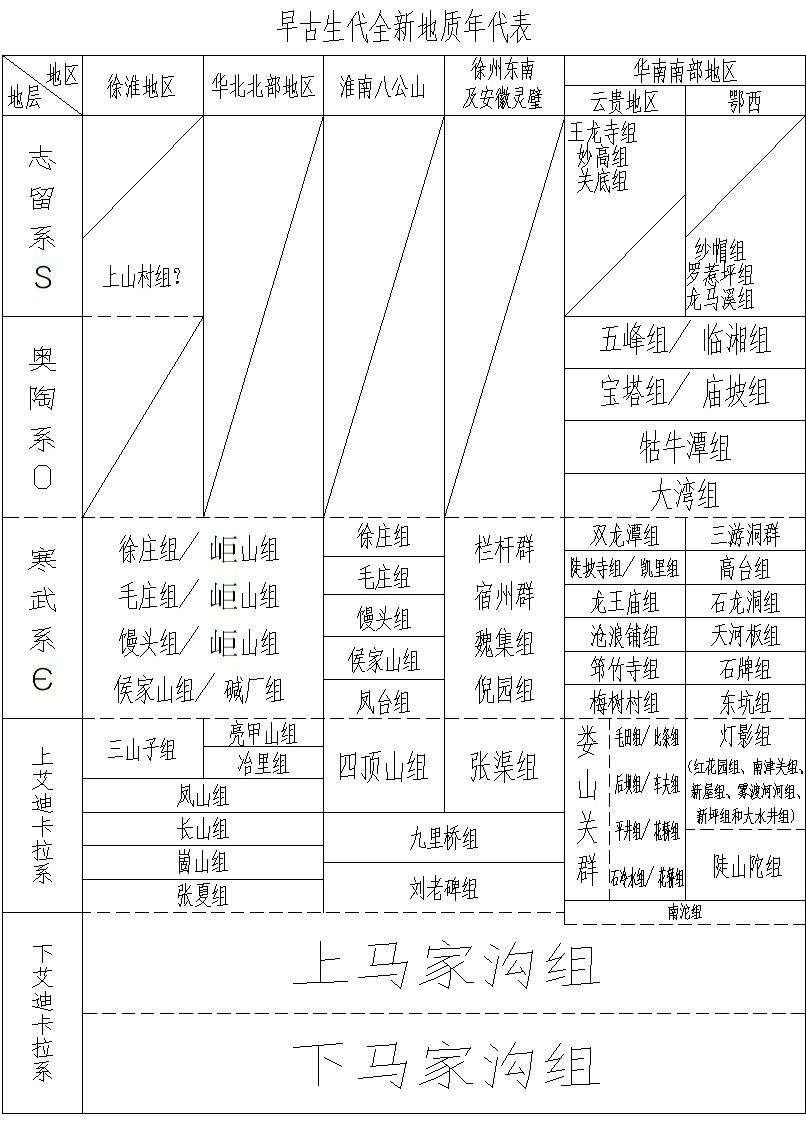
附笔者编制全新地质年代表
 九里山人 2016/8/8
九里山人 2016/8/8

评论
发表评论